NUOVE PROSPETTIVE SUL RUOLO DEI "VINCOLI"
(CONSTRAINS) NELL'EVOLUZIONE
Michele Sarà
Istituto di Zoologia
Via Balbi 5, 16126 Genova
e-mail zoologia@unige.it
Introduzione
Il corso e i risultati dell’evoluzione biologica sono regolati da "vincoli" (constraints) che agiscono dando limiti e forma ad ogni processo evolutivo. Perciò un interesse crescente è stato rivolto in anni recenti alla questione intricata del ruolo dei constraints in evoluzione (Gould, 1980, 1989; Alberch, 1982; Maynard Smith et al., 1985; Stearns, 1986; Wagner, 1988; Antonovics e Van Tienderen, 1991; Arnold, 1991; Mc Kitrick, 1993; Müller e Wagner, 1996).
Attualmente, la teoria dominante sui meccanismi evolutivi è la teoria neo-darwiniana o sintetica. Questa teoria è basata su di un meccanismo microevolutivo, prodotto da una sorgente casuale di variabilità genetica (mutazioni e ricombinazioni) e dal filtro della selezione naturale che sceglie fra fenotipi diversi. Naturalmente, la selezione naturale è efficace solo quando la diversità fenotipica può essere ereditata. Pertanto la scelta fra fenotipi diventa una scelta fra genotipi. Secondo l’ortodossia neo-darwiniana questo meccanismo è sufficiente per spiegare l’intero quadro evolutivo, compresa la macroevoluzione.
Tuttavia vari scienziati, specialmente nelle ultime due decadi, non concordano su queste posizioni. La selezione naturale viene considerata come più o meno importante ma non come il solo agente del processo evolutivo. Fondamento del neo-darwinismo è che la variabilità genetica non sia orientata ma del tutto casuale. Solo in questo caso infatti la selezione naturale può essere considerata unico agente costruttivo dell’evoluzione. Pertanto la sfida maggiore all’ortodossia neo-darwiniana viene dai fattori denominati "vincoli" (constraints), i quali operano non solo sul polo della selezione (vincoli selettivi) ma anche su quello della variabilità genetica (gli altri vincoli), direttamente o indirettamente attraverso la sua espressione nel fenotipo che è oltremodo complessa. Gould (1989), per esempio, indica i constraints come una fonte di cambiamenti o di restrizioni sui cambiamenti che non sorgono per selezione naturale e quindi operano al di fuori del quadro della teoria neo-darwiniana. I constraints possono derivare da una molteplicità di cause ma particolare attenzione - come polo opposto alla selezione naturale - è stata rivolta alle cause interne, vincoli formali che sorgono dalla base fisico-chimica delle strutture degli esseri viventi. Questi vincoli possono essere universali, riguardare cioè tutti i viventi oppure locali, dipendendo dalle caratteristiche chimiche e fisiche presenti in determinati taxa e organi (Maynard Smith et al., 1985). Essi sono legati ai constraints di sviluppo che operano nella costruzione del fenotipo.
Il peso dei constraints interni nel produrre i caratteri fenotipici e la plausibilità di un ruolo istruttivo dei fattori ambientali nelle loro genesi, e quindi nella genesi degli adattamenti, mediante meccanismi epigenetici mi hanno condotto (Sarà, 1993) a proporre un meccanismo evolutivo a due passi, epigenetico e selettivo, con una fase costruttiva epigenetica che si verifica nell’organismo e precede la fase selettiva che si verifica nella popolazione attraverso la scelta degli organismi più adatti. In effetti, l’argomento dei constraints, anche se molto ampio, intricato e confuso, può servire a comprendere meglio il pattern evolutivo che risulta da una rete intricata di molte cause interagenti.
Scopo di questo studio è di districare la confusione in termini e concetti attualmente esistente nell’argomento dei constraints e di dare un quadro più esauriente delle loro diverse categorie e del loro complicato intreccio.
Terminologia e concetti
È utile definire meglio il termine constraint ("vincolo"). In effetti, questo termine copre campi molto diversi di processi evolutivi ed è stato spesso adoperato in modo generico e confuso (Antonovics & Van Tienderen, 1991). Esiste una grande discrepanza su cosa debba essere inteso per constraint. Alcuni autori considerano come constraint evolutivo ogni causa (fattore, processo, situazione, condizione, ecc.) che determini un cambiamento in quanto dirige, "costringe" il cambiamento in un sentiero piuttosto che in un altro. Il cambiamento stesso rappresenta a sua volta un constraint. Pertanto i constraints sarebbero selettivi, quando prevale l’adattamento, di sviluppo quando è l’ontogenesi a dirigere e così avanti (Gould, 1989). Questo è l’uso che Arnold (1992) fa dei constraints genetici considerati come qualsiasi cambiamento che si verifica nel pattern della variazione e covariazione genetica. Pertanto, secondo questa veduta, la selezione direzionale è il risultato di constraints genetici.
Ma questo troppo largo, e pertanto generico, uso del termine constraint è stato criticato da Derrickson & Ricklefs (1988) quando essi sottolineano che un "constraint" filogenetico non è la stessa cosa di un "effetto" filogenetico, cioè della tendenza di organismi strettamente affini ad essere simili a causa della loro storia comune, una confusione fatta da molti biologi.
Sono pertanto d’accordo con Gould (1989) nel considerare il termine constraint come un termine relazionale che assume il suo significato solo in un contesto teorico, che è quello dettato dall’attuale dominante paradigma neo-darwiniano. Poichè in questo ogni cambiamento evolutivo si verifica attraverso selezione naturale operante sulla variabilità genetica casuale, i constraints sono secondo Gould (1989) "ciò che dirige altri tipi di cambiamenti o che impedisce cambiamenti che sarebbero operati dalla selezione". Pertanto Gould focalizza specialmente sui constraints di sviluppo che sono in larga parte influenzati dai constraints formali, cioè dalle regole strutturali dell’organismo, e dai constraints storici, cioè dalle contingenze della filogenesi. Il motivo di questa focalizzazione sta nel fatto che i constraints di sviluppo contrastano con una forma ristretta di funzionalismo darwiniano che, nella veduta di Gould, si è dimostrata inadeguata a spiegare, in particolare, i patterns ontogenetici e macroevolutivi.
Un altro punto importante da delucidare è il significato della parola constraint. Secondo Gould (1989) il termine constraint, dal latino stringere ha nella sua radice etimologica connotazioni sia positive che negative. Le implicazioni negative, cioè la nozione che i constraints siano solo limiti imposti al lavoro della selezione naturale, prevalgono largamente nell’uso del termine. Questo diminuisce il valore del loro impatto nell’evoluzione. Ma i constraints hanno anche un significato positivo. Ad esempio, nella definizione di Gould, i constraints sono fattori "che costringono o canalizzano i cambiamenti fenotipici in una direzione stabilita dalla storia passata o dalla struttura formale anzichè dal corrente adattamento" (che deriva dalla selezione naturale). Maynard Smith et al. (1985) considerano i constraints di sviluppo come una limitazione della variabilità fenotipica ma si domandano anche se "lo sviluppo si limiti a prevenire l’evoluzione dal seguire particolari sentieri oppure se essa serva anche come una forza direttiva, e risponda quindi parzialmente per gli aspetti orientati di vari trends e patterns". Secondo Alberch (1982) lo sviluppo può sfociare in discontinuità e direzionalità nelle trasformazioni morfologiche. Anche Riedl (1978), Bürger (1986) e Wagner (1988) sottolineano il ruolo attivo dei constraints. Associandomi ad essi darò risalto, nella successiva trattazione dell’argomento, a questo significato positivo del termine constraint. L’alternativa sarebbe di considerare la selezione naturale come il principale se non l’unico agente dell’evoluzione e i constraints solo come fattori di confine che condizionano il lavoro della selezione naturale. Questo significa non solo un’accettazione a priori dell’ortodossia neo-darwiniana ma contrasta anche con quanto emerge dallo studio dei constraints che anche se interagenti con la selezione si rivelano come vere e autonome forze evolutive. La relazione dei constraints con la selezione naturale è comunque lungi dall’essere chiara (Dullemeijer, 1991). In effetti, i soli constraints che sono presi in considerazione nel meccanismo evoluzionistico neo-darwiniano sono quelli denominati correntemente constraints selettivi.
Un’altra importante questione riguarda la definizione dei differenti tipi di constraints, attualmente confusa. Le categorie più utilizzate in letteratura sono quelle dei constraints di sviluppo, filogenetici, genetici e selettivi. I constraints di sviluppo sono basati, secondo Gould, su constraints formali e storici. I constraints filogenetici sono storici e in parte legati ai constraints di sviluppo.
Si è detto che i constraints genetici sono stati definiti da Arnold (1992) come "qualsiasi cambiamento nel pattern della variazione e covariazione genetica per una serie di caratteri". Ma questi cambiamenti, come indicato dallo stesso Arnold, sono essenzialmente l’effetto della selezione naturale: pertanto, a mio avviso, essi non si devono considerare constraints ma conseguenze "canoniche" della teoria neo-darwiniana. Un esempio di veri constraints genetici, non direttamente legati alla selezione naturale, sono i cambiamenti del genoma dovuti agli elementi trasponibili.
Anche l’uso dei constraints selettivi è controverso. Arnold applica il termine ad ogni modificazione della selezione naturale che alteri il pattern della variazione e covariazione genetica. Anche Amundson (1994) considera i constraints selettivi come constraints sulla variabilità genetica casuale, da lui indicata come drift. Maynard Smith et al. (1985) parlano di constraints selettivi come constraints che la selezione opera sullo sviluppo per formare gli adattamenti. In tutti questi casi i constraints selettivi sono identificati con lo stesso operare della selezione naturale quale selezione direzionale.
In coerenza con l’uso del termine constraint nelle altre categorie penso invece che il termine di constraints selettivi debba essere applicato a quei fattori, soprattutto stocastici, che alterano i risultati attesi della selezione, come il drift (deriva genetica), effetto collo di bottiglia, isolamento, ecc. Così intesi i constraints selettivi sono constraints in quanto alterano le previsioni del meccanismo selettivo di base, dato dai valori di fitness. Essi, diversamente dagli altri constraints, sono presi in debita considerazione dalla teoria neo-darwiniana. Antonovics & Van Tienderen (1991) giungono alla conclusione che la necessità di usare il termine constraint possa essere completamente ovviata, in quanto i constraints sarebbero compresi entro il paradigma della teoria sintetica. Ma essi si riferiscono a quelli che io definisco constraints selettivi in quanto condizioni che modificano il loro "modello nullo", cioè il corso predicibile dell’evoluzione basato sulla selezione naturale, cioè sui valori di fitness, e non prendono in considerazione gli altri constraints.
Come modifiche al "modello nullo" sono citati da questi autori: lo spettro delle mutazioni/ricombinazioni a disposizione, lo squilibrio pleiotropia/associazione fra i caratteri, la struttura di popolazione condizionata da piccole dimensioni di popolazione, flusso genico, dalla selezione a livelli non individuali o anche da risposte coevolutive di specie associate. Comunque la fissazione di mutazioni neutrali o la speciazione attraverso effetto del fondatore sono previsti nella teoria sintetica. Il fatto che Antonovics e Van Tienderen (1991) si limitino a quelli che io definisco constraints selettivi è soprattutto dovuto al loro pensare "genetico-popolazionistico" che dà a a priori alla teoria della selezione naturale il ruolo di "ipotesi nulla". Pertanto essi tralasciano gli altri constraints, i non selettivi, come quelli filogenetici, formali, di sviluppo. Questi sono fattori che modificano il corso evolutivo, indipendentemente dai cambiamenti nella selezione naturale. Essi non dirigono o modulano la selezione anche se interagiscono con essa.
Antonovics & Van Tienderen (1991), quale commento alle asserzioni di Gould (1989), dicono che essi preferiscono il termine "ipotesi nulla" a "teoria favorita", come usato da Gould, perchè quest’ultimo è pregiudizievole e suggerisce implicitamente che la teoria favorita sia inadeguata. In realtà, Gould, asserendo che vada fatta una distinzione fra una teoria favorita e i constraints esterni a questa teoria, prende una posizione non prevenuta verso qualsiasi teoria evolutiva. All’opposto, Antonovics & Van Tienderen, considerando quali constraints della loro "ipotesi nulla" solo quelli selettivi, che sono interni alla teoria neo-darwiniana perchè riguardano il suo fattore centrale, la selezione naturale, partono da una posizione pregiudiziale neo-darwiniana, che quindi ignora le altre autonome categorie di constraints.
Invece, le categorie autonome, "esterne" di constraints sono numerose e spesso non meno importanti della selezione naturale e dei constraints selettivi per il processo evolutivo. Nei paragrafi seguenti discuterò le varie categorie di constraints normalmente prese in considerazione. Introdurrò anche due nuove categorie, i constraints epigenetici e di comportamento, rese necessarie dal progresso della ricerca evoluzionistica.
Un altro punto che non è stato preso finora in sufficiente considerazione riguarda l’interazione fra le diverse categorie di constraints e fra queste e la selezione naturale. Il ragionamento di Amundson (1994) sul ruolo dei constraints di sviluppo negli adattamenti in cui vengono contrapposte nettamente una visuale adattazionista selettiva e una visuale "di sviluppo" deriva dal non aver preso in considerazione che, pur se fra fattori autonomi, fra selezione naturale e constraints di sviluppo deve necessariamente, nel processo evolutivo, stabilirsi un’interazione.
I constraints formano nel loro insieme un’intricata rete di cause con influenze reciproche bidirezionali. I constraints, siano essi più condizionati dal sistema genetico o dall’ambiente, non possono essere considerati separatamente, perchè formano una rete, i cui elementi differiscono solo, nell’influenzare un processo evolutivo, per la loro relativa importanza (Dullemeijer e Zweers, 1997). Questi autori considerano l’analisi della rete di constraints come un problema dei tre corpi allargato, un problema multirelazionale, che non può essere ridotto da successive analisi delle connessioni perchè ogni relazione ha un effetto simultaneo sulle altre relazioni. Il sistema di constraints evolutivi è pertanto indeterminato (Prigogine e Stenger, 1997). Nella ricerca evolutiva ciò è ulteriormente complicato dal fatto che storicamente non tutti i cambiamenti hanno lo stesso effetto. Piccoli cambiamenti in alcuni punti del sistema possono avere grandi effetti sull’intera rete d’interazioni mentre grandi cambiamenti in altri posti possono agire poco o nulla. Ciò è connesso al problema delle innovazioni chiave, quelle che determinano effetti evolutivi a lungo termine. Il problema dei constraints è ulteriormente complicato dalle loro interazioni con la selezione naturale. La constatazione che il pattern dei constraints è reticolato porta ad una visione più olistica del processo evolutivo.
Le categorie di constraints
a) Constraints strutturali
I constraints strutturali rappresentano una vasta categoria di vincoli interni. Questo
nuovo termine comprende i constraints "universali" e alcuni constraints
"locali" sensu Maynard Smith et al. (1985). I constraints universali si
applicano a tutti i sistemi fisici, compresi gli organismi, costruiti con un dato
materiale o provvisti di una data complessità. Un vincolo universale è, per esempio,
rappresentato dalla tensegrità, la regola generale per l’autoassemblaggio delle
strutture biologiche (Ingber, 1993, 1997). I vincoli strutturali che dipendono dalle
proprietà degli organismi come conseguenza fisica della loro immediata struttura sono
sono stati chiamati da Gould (1989) constraints formali. Essi corrispondono ai
"meccanismi generici" di Newman e Comper (1990). I vincoli strutturali non sono
soltanto universali ma anche locali, dipendendo dal particolare materiale utilizzato in un
dato taxon o organo. Il confine fra i vincoli universali e locali, come osservato da
Maynard Smith et al. (1985), non è del tutto chiaro e neppure del tutto rigido. Essi
formano un continuum e può anche essere riconosciuta una categoria intermedia di vincoli
"generali". Per Alexander (1985) i vincoli universali sono basati sulle
proprietà universali della materia e dell’energia e gli organismi possono fare uso
solo di una parte limitata di questo spettro di proprietà. I vincoli generali sono quelli
che dipendono dai tipi di materiale e dai principi costruttivi mentre i vincoli locali
sono dati dall’interazione di strutture. Ma i termini universale, generale e locale
sono utilizzati anche in altro modo. Per questo motivo preferisco raggruppare tutti i
constraints, basati sulle leggi della fisica, sui materiali mediante cui gli organismi
sono costruiti e sulle loro caratteristiche matematicamente derivabili di sistemi
complessi, nella categoria dei constraints strutturali, indipendentemente dalla loro
ricorrenza più o meno universale. I vincoli legati alle proprietà dei sistemi complessi
sono stati anche chiamati constraints topologici (Kauffman, 1985).
Un rapporto stretto esiste fra i constraints strutturali, soprattutto quelli locali, e i constraints di sviluppo. Un altro rapporto esiste fra i constraints strutturali e le loro capacità funzionali. Focalizzando su queste capacità, alcuni vincoli strutturali sono stati chiamati constraints funzionali o "design constraints" (Lauder, 1996) per la loro interazione con il processo di selezione naturale. I constraints strutturali rinforzati da richieste funzionali possono influenzare fortemente il pattern macroevolutivo e filogenetico (Carrier, 1991; Herring, 1993).
b) Constraints di sviluppo
I constraints di sviluppo rappresentano la categoria di vincoli maggiormente presa in
considerazione quale elemento di contrasto alla selezione naturale (Maynard Smith et al.,
1985). L’uso di questo termine si riferisce ad una categoria molto composita. Molti
constraints di sviluppo sono la conseguenza di constraints strutturali. Per esempio, molte
caratteristiche del sistema nervoso centrale dei mammiferi possono essere spiegate
mediante un meccanismo morfogenetico che coinvolge una tensione meccanica lungo gli
assoni, i dendriti e i processi della glia. Nella corteccia cerebrale la tensione lungo
gli assoni della materia bianca può spiegare come e perchè la corteccia si piega in un
caratteristico pattern specie-specifico. (Van Essen, 1997). Ma, oltre che da proprietà
dei materiali di costruzione, i constraints di sviluppo possono sorgere anche (Maynard
Smith et al., 1985) da richieste concernenti l’immagazzinamento e il riutilizzo
dell’informazione impiegata nello sviluppo. Questo può essere dovuto alle
particolari caratteristiche delle vie di sviluppo evolutivamente determinate di un dato
gruppo di organismi o alle caratteristiche matematiche di una classe di sistemi entro cui
si situa un dato sistema di sviluppo. Una visione auto-organizzativa dei constraints di
sviluppo, non innescati da composizione molecolare o da esigenze funzionali, è stata
tracciata da Berrill e Goodwin (1996). I vincoli di sviluppo sono allora prodotti dalla
dinamica delle reti genetiche che agiscono nel contesto dei campi morfogenetici e sono
quindi il risultato di processi interattivi di auto-organizzazione. La nozione di campi
morfogenetici è pertanto centrale in questa visuale strutturalista dei vincoli di
sviluppo.
I constraints di sviluppo, che derivano spesso ma non sempre dai constraints strutturali, possono sorgere anche a causa di contingenze storiche, e quindi dai constraints filogenetici. D’altra parte essi rappresentano la fonte dei constraints filogenetici e quindi di cambiamenti nel pattern macroevolutivo. Maynard Smith et al. (1985) danno la seguente definizione non-convenzionale dei constraints di sviluppo: "un constraint di sviluppo è un’anomalia nella produzione dei fenotipi varianti o una limitazione nella variabilità fenotipica causata dalla struttura, carattere, composizione o dinamica del sistema di sviluppo". Questa definizione è coerente con il fatto che i constraints di sviluppo non funzionano nel secondo stadio ecologico del meccanismo neo-darwiniano, il processo di selezione naturale, ma nel primo stadio, la produzione di variazioni ereditabili. Pertanto, come suggerito da Amundson (1994), i constraints di sviluppo non agiscono sull’adattamento, come sostenuto da Stephen e Krebs (1986) ma sulla forma organica come indicato da Alberch (1992). Solo indirettamente un vincolo sulla forma può diventare un vincolo sull’adattamento. Questa visuale sui constraints di sviluppo sottolinea che non è sufficiente conoscere i genotipi che determinano i fenotipi ma occorre anche conoscere le interazioni causali che legano genotipi e fenotipi.
I constraints di sviluppo interessano un largo spettro di fenomeni biologici, dai cambiamenti eterocronici nello sviluppo a mutamenti nella plasticità e anche nelle storie vitali che possono influenzare profondamente il pattern macroevolutivo (Smith Gill, 1983; West-Eberhard, 1989). I vincoli di sviluppo sono considerati come un importante fattore per mantenere cicli vitali complessi (Moran, 1994), promuovendo un disaccoppiamento adattativo che favorisce l’indipendenza dei caratteri larvali e adulti. D’altra parte, i constraints di sviluppo possono anche mantenere condizioni stabili come lo stadio filotipico degli animali. Esse sono state prodotte dai constraints genetici responsabili, insieme con le forze selettive, della formazione dei geni homeobox. A sua volta, la presenza di stadi filotipici determina constraints filogenetici nel corso macroevolutivo.
c) Constraints filogenetici
Denominati anche storici, i constraints filogenetici possono essere definiti secondo
McKitrick (1993) come "qualsiasi risultato o componente della storia filogenetica di
un lignaggio che altera il previsto corso evolutivo di quel lignaggio".
I constraints filogenetici sono strettamente legati ai constraints di sviluppo perchè, come mostrato da Gould (1989), l’ontogenesi è il mezzo attraverso cui i constraints storici sono espressi. I constraints di sviluppo che determinano i constraints filogenetici sono quelli chiamati da Maynard Smith et al. (1995) come "locali" perchè si applicano ad uno spettro limitato di taxa. Essi influenzano la storia evolutiva impedendo ad una specie o gruppo di specie di seguire alcune vie evolutive. Nella storia della vita i piani strutturali degli organismi possono essere conservati o cambiati. La conservazione di un piano può essere dovuta a un constraint attivo di sviluppo dipendente dalle necessità di funzionamento. All’inverso, la comparsa di una novità chiave può determinare la liberazione da un precedente vincolo di sviluppo e filogenetico. La novità chiave diventa un nuovo constraint che promuove l’innovazione invece della conservazione, favorendo la radiazione adattativa. Pertanto, la successione dei constraints filogenetici, anche di segno opposto, modella il pattern macroevolutivo.
Inoltre, i constraints filogenetici sono molto importanti nella storia della vita perchè, in una modalità più stocastica, possono dipendere da eventi imprevedibili come estinzioni di massa o onde di migrazione, legate alla storia paleoecologica e geologica della terra, come si verificò alle transizioni Permiano-Triassico (P-T) e Cretacico-Terziario (C-T). Improvvise fluttuazioni climatiche, dovute, ad esempio, alla caduta di grandi meteoriti, variazioni nel livello dei mari e nel profilo dei continenti, possono provocare rapidi e generali cambiamenti dei biota. In tal modo i constraints filogenetici hanno influenzato profondamente, con restrizioni e scomparsa di talune linee filetiche ma anche con comparsa ed espansioni di altre l’evoluzione degli organismi al di là dell’azione gradualistica della selezione naturale.
d) Constraints genetici
I constraints genetici riguardano alterazioni nel meccanismo genetico stesso. Come
osservato da Maynard Smith et al. (1985) vi è un accumulo continuo nell’evoluzione
di nuovo materiale genetico con aumento del contenuto in DNA della cellula e del numero di
cromosomi. Questo meccanismo di accumulo rappresenta in se stesso un constraint e una
delle sue più importanti fonti, oltre alle duplicazioni di geni, è data dalle
modificazioni del genoma dovute al sistema di trasposoni (Van Sternberg, 1996). I
trasposoni partecipano allo sviluppo del genoma perchè sono cooptati in ruoli strutturali
e regolatori. Essi non agiscono come elementi genetici autonomi ma vengono vincolati da
effetti di posizione e dalle reti regolatrici del genoma dell’ospite. Essi entrano
nel processo di evoluzione concertata, conosciuto come "deriva molecolare"
(Dover, 1982) in cui la dispersione degli elementi di una sequenza "favorita" si
verifica attraverso i meccanismi di una conversione genica alterata, di scambi ineguali
fra cromatidi fratelli e di trasposizione duplicativa.
Altri fenomeni genetici in cui il genoma si auto-trasforma sono le paramutazioni, note solo per le piante, e il silenziamento dei geni, ottenuto anche nella sperimentazione con transgeni. Nelle paramutazioni la struttura cromatinica di un allele può alterare lo stato cromatinico del suo omologo o paralogo in un modo direttamente ereditabile (Brink, 1960; Mikula, 1995). Il silenziamento transgenico è affine alla paramutazione nel suo meccanismo ma non è limitato ad alleli o a cromosomi omologhi (Jorgensen, 1994). Paramutazione e silenziamento transgenico possono essere inclusi in una nuova categoria di vincoli, trattata in seguito, che può essere denominata dei constraints epigenetici e a cui appartengono anche le epimutazioni (Holliday, 1987; Jablonka e Lamb, 1995). Infatti, l’effetto sul corso evolutivo di paramutazioni e silenziamento transgenico è, come per le epimutazioni, dovuto ad un cambiamento nell’espressione genica che induce cambiamenti fenotipici. Van Sternberg et al. (1992) considerano le auto-modificazioni del genoma come una serie ricorrente di cambiamenti di stato influenzati e diretti che si verificano dai batteri all’uomo.
Un altro aspetto dell’autoregolazione del genoma è dato dalla duplicazione genica. Questa permette l’emergenza di nuovi geni con nuove funzioni. L’auto-organizzazione del genoma porta i genomi a individualizzarsi, svilupparsi e crescere in complicazione accompagnandosi a compartimentalizzazione. Questo processo non sembra dovuto a selezione ma a vincoli genetici.
I constraints genetici possono esser stimolati dall’ambiente interno, cellulare, e da quello esterno. A mio avviso la più importante linea di ricerca sui constraints genetici prodotti da stimoli ambientali dovrebbe essere rappresentata dal meccanismo di assimilazione genetica, un processo che ha la sua base teoretica nell’effetto Baldwin (1896) ed è stata dimostrata sperimentalmente dalla ricerca di Waddington (1957, 1975). Il processo di assimilazione genetica è ben stabilito, anche se su pochi casi, ma la sua interpretazione è molto discussa. È verosimile che il processo di assimilazione genetica non sia ristretto a singoli casi ma si verifichi frequentemente soprattutto per i caratteri comportamentali (Tierney, 1986; Vancassel, 1990; Gerard et al., 1992). Un fenomeno collegato è quello dell’esistenza delle fenocopie (Kauffman, 1993 ); inoltre l’assimilazione genetica consentirebbe ad una parte della plasticità fenotipica di divenire ereditaria.
e) Constraints epigenetici
Si tratta di una nuova categoria di constraints la cui introduzione è richiesta dai
recenti sviluppi della biologia molecolare e delle conoscenze nel funzionamento della
cellula. Si tratta anche dei constraints più importanti nel trasferire direttamente nella
realizzazione del fenotipo l’informazione ambientale.
La corrente teoria adattazionista basata solo sul processo della selezione naturale è, a mio avviso, contraddetta dal fatto che il lavoro dei constraints di sviluppo entro i campi morfogenetici è aperto all’influenza diretta dell’ambiente esterno ed interno. Questa nuova categoria di constraints, detti epigenetici, rappresenta la cinghia di trasmissione fra l’ambiente, il sistema genetico e i campi morfogenetici, entro cui si formano i constraints di sviluppo. Essi rappresentano pertanto per l’organismo una seconda fonte d’informazione ambientale in aggiunta alla fonte selettiva.
"Epigenetico" è il termine correntemente usato dalla genetica e citologia molecolare per ogni processo in cui l’azione dei geni sia promossa, regolata o repressa da agenti citoplasmatici o esterni. Il ruolo degli agenti epigenetici e degli annessi sistemi epigenetici sembra molto ampio perchè riguarderebbe non solo l’espressione e regolazione genica nei fenomeni di trascrizione ma anche i processi di riparo del DNA e di mutazione. Il loro impatto evolutivo è rafforzato dal fatto che agenti epigenetici possono essere ereditati attraverso sistemi di eredità epigenetica (EIS) (Jablonka e Lamb, 1995). I constraints epigenetici pssono essere definiti come ogni anomalia nella produzione di fenotipi varianti causata dal carattere e dalla dinamica degli agenti epigenetici.
I vincoli epigenetici più studiati sono quelli dovuti alla metilazione del DNA che determinano repressione o silenziamento genico nell’imprinting genomico; essa implica un’espressione differenziale dei geni paterni e materni (Monk, 1995).
Effetti parentali non-mendeliani (comprendenti gli effetti materni) sono un’altra fonte di fattori che possono influenzare o regolare l’espressione di alleli. Essi inducono cambiamenti fenotipici nella prole (Carrière, 1994), possono produrre effetti ambientali indiretti (IEE) (Rossiter, 1996) e sono stati considerati di natura ambientale anche nel quadro della genetica quantitativa (Kirkpatrick e Lande, 1989). Questi effetti parentali possono influenzare la relazione fra i fenotipi degli individui di una famiglia e i loro valori di fertilità, modificando la covarianza genetica fra i caratteri espressi in ambienti diversi o introducendo ritardi di tempo nella loro risposta alla selezione. Come mostrato nella documentata sintesi di Rossiter (1996) gli effetti ambientali ereditati possono avere un vasto impatto sulla dinamica di popolazione, sull’espressione della plasticità fenotipica adattativa e quindi sull’evoluzione dei caratteri. Un esempio è dato da condizioni ambientali, quali quelle fotoperiodiche, climatiche o alimentari, che, sperimentate nella generazione parentale, possono influenzare il fenotipo della prole e delle generazioni successive con un impatto sui caratteri delle storie vitali e sulla loro ereditarietà.
Successivamente, Wolf et al. (1998) hanno mostrato che le influenze ambientali sul fenotipo di un individuo possono essere dovute all’espressione dei geni in un differente individuo conspecifico, che può essere un parente o anche un individuo non imparentato e non necessariamente un genitore. Essi chiamano questo fenomeno effetti genetici indiretti (IGE). Gli IGE portano ad un pattern complicato di eredità, in cui le sorgenti ambientali di variazione possono essere trasmesse attraverso le generazioni e quindi contribuire ai cambiamenti evolutivi. L’esistenza degli IGE altera la relazione fra fenotipo e genotipo e pertanto produce un effetto epigenetico che modifica il corso dell’evoluzione, nelle parole di Wolf et al., "in un modo drammatico e non-intuitivo". In questo processo gli stessi ambienti, come effetti ambientali indiretti (IEE) che riflettono gli effetti indiretti sui geni, sono potenzialmente sottoposti a selezione e conseguente evoluzione. L’importanza degli IGE deriva dal fatto che essi non riguardano solo gli effetti materni ma anche altre interazioni fra individui e influenzano caratteri di vario tipo dallo sviluppo al comportamento, ad esempio in specie con cure parentali o dispersione differita. Gli IGE possono essere alla fonte degli effetti degli ambienti sociali sul comportamento in modalità come la comunicazione, l’accoppiamento, l’aggressività e l’approvvigionamento, e pertanto anche sui constraints di comportamento (Wolf et al., 1998).
Il ruolo evolutivo degli IGE può essere legato in un contesto più ampio con i problemi della sensitività allelica e della regolazione genica della plasticità fenotipica (Pigliucci e Schlitchting, 1997). In entrambi questi fenomeni vi è una percezione diretta dell’ambiente da parte del sistema genetico e questa percezione è tradotta in effetti fenotipici. Successivamente il pattern fenotipico così cambiato in una generazione è sottoposto alla selezione naturale, che quindi agisce non solo su un pattern modificato di genotipi ma anche, attraverso il nuovo pattern, sulle condizioni ambientali a cui essi furono sensitivi. Ciò è coerente con un meccanismo evolutivo a due stadi, epigenetico e selettivo, come da me precedentemente suggerito (Sarà, 1993).
Altri constraints epigenetici, che implicano un’espressione genetica influenzata dalle condizioni ambientali esterne ed interne, sono dati dalle epimutazioni, paramutazioni e dal silenziamento transgenico (paramutazioni e silenziamento transgenico possono essere inclusi anche fra i constraints genetici). Anche l’epistasi, cioè l’effetto non-additivo dell’interazione genetica, è una sorgente di forti effetti fenotipici e quindi di vincoli epigenetici. La frequenza di tali effetti può aumentare in presenza di fattori che influenzano le frequenze genotipiche in una popolazione quali la selezione direzionale, l’inincrocio o i colli di bottiglia, alcuni dei quali possono essere considerati come vincoli selettivi. L’epistasi è importante al livello fenotipico perchè può influenzare i coefficienti di selezione a cui sono sottoposti gli alleli e pertanto modificare i ritmi evolutivi, con un ruolo creativo nell’evoluzione (Moreno, 1994).
I constraints epigenetici possono agire sullo sviluppo producendo constraints di sviluppo. Per esempio, come suggerito da Müller e Wagner (1996), è verosimile che la transizione pinna /arto nei vertebrati sia stata causata da cambiamenti epigenetici nel contesto dell’azione dei geni Hox. In questo caso il cambiamento evolutivo sembra aver riguardato in primo luogo la durata della proliferazione mesenchimatica anzichè lo stesso codice degli Hox. In effetti, chiari esempi di effetti epigenetici possono essere trovati anche in altri tessuti connettivi. Questi sistemi reagiscono fortemente alle influenze fisiche nello sviluppo embrionale, quali lo stress meccanico (Hennig, 1993). Le forze di pressione, per esempio, possono provocare la formazione di cartilagine secondaria o di osso di membrana (Hall, 1986).
Processi come lo "splicing" (montaggio) dell’RNA, la trasduzione dei segnali e i patterns di formazione dei tessuti e degli organi sono processi informazionali di base nella costruzione della forma corporea, allo stesso modo della sintesi proteica ad opera del codice genetico del DNA. Essi non sono dovuti a proprietà intrinseche, strutturali o funzionali, come i constraints strutturali o di sviluppo ma, secondo Barbieri (1997), a codici convenzionali per la trasmissione dell’informazione addizionali al codice genetico del DNA. I constraints genetici, come quelli legati agli elementi trasponibili, sono constraints informazionali ma anche i suddetti processi, in gran parte epigenetici, portano a modifica nell’informazione ereditaria, e il termine di constraints informazionali può applicarsi anche ad essi. Pertanto si potrebbe istituire una categoria a sè di constraints informazionali comprendente processi di natura diversa. Ma le conoscenze su questi processi informazionali e sul loro funzionamento nella cellula non sono ancora approfondite. Ad esempio, per Barbieri occorre distinguere fra processi informatici e semantici (portatori di significato), questi ultimi legati a codici. Ho preferito in questa trattazione includere tali vincoli fra i constraints epigenetici che raggruppano tutti i processi che forniscono all’organismo un’informazione supplementare a quella data dal sistema genetico.
f) Constraints comportamentali
Viene proposta questa nuova categoria di constraints per cambiamenti nelle relazioni
con l’habitat (comprendenti anche la scelta degli habitat). Esse possono essere
dovute sia a modifiche nei meccanismi dell’istinto e quindi nei relativi constraints
di sviluppo sia a processi di apprendimento. Ad esempio, la scelta dell’habitat da
parte d’individui o di popolazioni può innescare una cascata di eventi che produce
effetti fenotipici diretti attraverso la plasticità fenotipica e non solo indirettamente
attraverso la selezione naturale.
Un ruolo speciale è comunque assunto dall’apprendimento in quanto legato alla plasticità neurale. In questo tipo di comportamento la relazione con i geni diventa molto tenue ed indiretta. L’apprendimento è fortemente dipendente dalle influenze ambientali e diventa una sorgente di constraints di comportamento che contrastano con la selezione naturale e influenzano la plasticità fenotipica. Esso rappresenta pertanto un canale attraverso cui constraints epigenetici (Avital e Jablonka, 1994) ma anche constraints genetici grazie a processi di assimilazione genetica (Tierney, 1986) possono essere provocati nell’organismo. D’altra parte constraints epigenetici, come descritto prima per gli IGE, possono produrre constraints di comportamento.
Gli effetti coevolutivi nella predazione, parassitismo e simbiosi possono provocare sia constraints di comportamento che epigenetici. L’associazione fra specie determina in entrambi i partners cambiamenti nel comportamento ma anche cambiamenti nei meccanismi epigenetici che controllano la morfogenesi e lo sviluppo.
g) Constraints ecologici o ambientali.
Questa è una categoria di vincoli composita e mal definita. Ritengo che, almeno in larga
parte, i cosiddetti constraints ecologici possano essere inclusi nelle categorie dei
vincoli di comportamento ed epigenetici, che rappresentano l’effetto
sull’organismo delle influenze ambientali dirette. I constraints ecologici che
secondo Faulkes et al. (1997) guidano l’evoluzione sociale negli eterocefali ciechi
africani, sono da considerarsi, nella prospettiva di questo articolo, come constraints di
comportamento. Componenti ecologiche esistono anche nei constraints filogenetici (ad
esempio i grandi cambiamenti climatici) e in quelli di sviluppo (ad esempio le richieste
funzionali). Il processo ecologico fondamentale che porta agli adattamenti delle
popolazioni, con cui vengono filtrati gli organismi a seconda della loro fitness, non è
ovviamente un constraint ecologico perchè è il processo stesso di selezione naturale.
Tuttavia, potrebbe essere opportuno definire una categoria di constraints ecologici, denominandola ecosistemici o biocenotici per taluni effetti evolutivi delle relazioni fra organismi negli ecosistemi. Queste relazioni non sono infatti solo di predatore-preda e pertanto regolate dalla selezione naturale ma anche basate su interazioni fra organismi dal parassitismo alla simbiosi. Soprattutto in quelle cooperative possono entrare in gioco altri fattori (constraints ecosistemici o biocenotici) che si legano a constraints comportamentali, epigenetici e di sviluppo e portano anche a constraints filogenetici.
h) Constraints selettivi
Raggruppo in questa categoria i fattori, essenzialmente stocastici e contingenti, che
alterano il corso della selezione naturale. Essi sono presi in considerazione nel
meccanismo di genetica delle popolazioni che rappresenta la base della teoria
neo-darwiniana. Fra questi constraints vi sono il "drift", i
"bottlenecks", l’isolamento, la migrazione, la densità e grandezza di
popolazione, ecc.
La rete dei constraints
I caratteri fenotipici morfologici, fisiologici e comportamentali dell’organismo sono sottoposti ad un’ampia e disparata serie di constraints evolutivi che si aggiungono all’azione della selezione naturale e interagiscono con essa. Ho riconosciuto e definito in questo lavoro le seguenti categorie di constraints: strutturali, di sviluppo, filogenetici, genetici, epigenetici, comportamentali e selettivi. Una categoria di constraints "ecosistemici" al posto dei constraints ecologici può anche essere individuata. I constraints, eccetto quelli selettivi, che sono incorporati nella teoria neo-darwiniana, contrastano e alterano, spesso in modo drammatico, gli esiti dell’evoluzione fenotipica previsti dall’azione reciproca della selezione naturale e della variabilità genetica casuale. Molti di questi constraints sono influenzati direttamente dalle condizioni ambientali e rappresentano quindi una seconda fonte evolutiva, in aggiunta alla selezione naturale, per l’adattamento degli organismi all’ambiente.
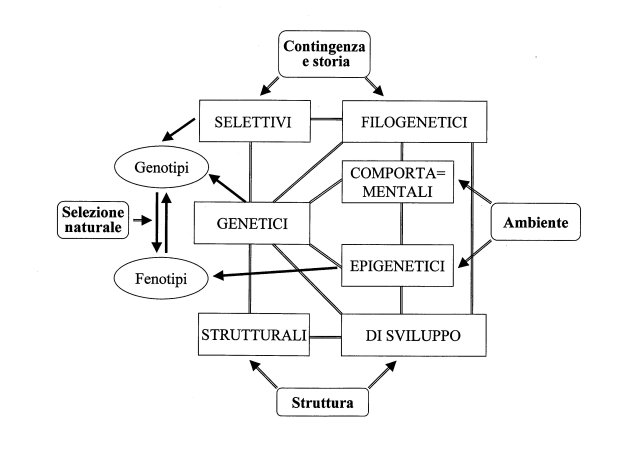
| fig. 1 - Rete dei vincoli (constraints) dell’evoluzione con le loro sorgenti (struttura, ambiente e contingenza-storia). La selezione naturale interagisce con i constraints modificando le frequenze dei genotipi/fenotipi. Nuovi fenotipi ereditabili possono essere prodotti non solo attraverso i genotipi ma anche attraverso i constraints epigenetici. |
I constraints non operano separatamente ma interagiscono fra loro e con la selezione naturale. L’evoluzione di ogni carattere di una specie e dell’organismo nel suo complesso, è dovuta alla rete intricata fatta dai constraints e dai constraints con la selezione. La rete dei constraints è prodotta da tre elementi: struttura, influenza dell’ambiente e storia o contingenza che integrano il lavoro della selezione naturale nel generare il pattern evolutivo. Questa rete (fig. 1) costituisce la base olistica del processo evolutivo, cioè il substrato biologico per l’evoluzione degli esseri viventi come interi e non come riunione di parti separate.
Diverse cascate, in gran parte bidirezionali, di effetti evolutivi sono individuabili in questa rete. Per esempio, i vincoli strutturali possono influenzare lo sviluppo, producendo vincoli di sviluppo, che determinano vincoli filogenetici ma quest’ultimi, a loro volta, possono produrre constraints strutturali e di sviluppo. D’altra parte l’ambiente può indurre vincoli comportamentali ed epigenetici e questi produrre vincoli di sviluppo e genetici. L’esistenza dei constraints accanto alla selezione naturale produce nel processo evolutivo una polarità fra:
a) un insieme di fattori costruttivi del fenotipo che comprende i constraints strutturali, di sviluppo, epigenetici, comportamentali, genetici (in parte) e filogenetici (in parte). Essi agiscono soprattutto a livello dell’organismo.
b) un insieme di fattori selettivi e storico-stocastici che comprende la selezione naturale, i constraints selettivi e in parte quelli genetici e filogenetici. Essi agiscono soprattutto a livello della popolazione.
I constraints genetici e filogenetici rappresentano quindi un’area di sovrapposizione fra i due insiemi (fig. 2).
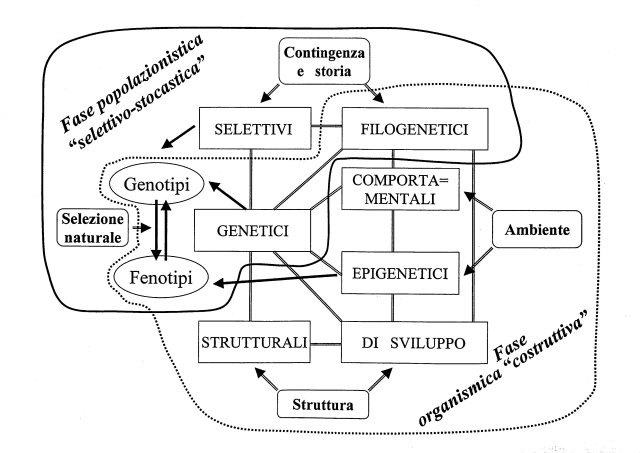
| fig. 2 - Pattern dell’evoluzione che include selezione naturale e vincoli (constraints). Esso è suddiviso in un insieme "costruttivo" che opera a livello di organismo e in uno "selettivo-stocastico" che opera a livello di popolazione. I constraints genetici, filogenetici e la produzione di genotipi/fenotipi si trovano in un’area di sovrapposizione. |
I due insiemi si riferiscono rispettivamente alle due fasi del meccanismo neo-darwiniano espresse: a) dalla produzione della variabilità ereditaria; b) dalla selezione naturale (con l’aggiunta del drift e di altri fattori storico-stocastici).
La considerazione dei constraints amplifica il polo della produzione della variabilità ereditaria mostrando che il fenotipo non è nella sua realizzazione un semplice specchio delle mutazioni e ricombinazioni geniche casuali.
L’evoluzione del fenotipo dell’organismo è influenzata da due elementi, ambiente e struttura, che agiscono come poli di un continuum. Essi generano con interazioni multiple bidirezionali constraints fondamentalmente costruttivi, come quelli strutturali, di sviluppo, epigenetici e di comportamento. La loro azione evolutiva si lega naturalmente al genoma sia per la necessaria base di memoria genetica sia per il passaggio delle informazioni, che possono però essere trasmesse anche per via epigenetica, nelle generazioni successive. Per la formazione degli adattamenti si deve naturalmente considerare l’interazione con la selezione naturale.
Correntemente, ad eccezione dei constraints selettivi, i constraints sono considerati, dai seguaci della teoria neo-darwiniana, come fattori limitanti o di confine di trascurabile o secondaria importanza nell’evoluzione. La mia proposta, suffragata dal crescente interesse nel ruolo evolutivo dei constraints, è che l’evoluzione sia diretta olre che dalla selezione naturale, dall’azione dei constraints, uniti in una rete.
Il mio modello (Sarà, 1993) di un meccanismo evolutivo a due passi che allarga il paradigma corrente della Sintesi riflette la polarità sopra indicata. I due passi si alternano e interagiscono ad ogni generazione. Il primo passo, costruttivo, organismico, è rappresentato essenzialmente dalla rete dei constraints (eccetto quelli selettivi). Il secondo passo, selettivo, popolazionistico, è dato dal processo di selezione naturale e dai constraints selettivi.
Ringraziamenti
Sono grato ai professori Elda Gaino e Giorgio Bavestrello per l’utile discussione e l’aiuto nella stesura del manoscritto e delle figure.
Ricerche effettuate con fondi del Ministero della Ricerca Scientifica e Tecnologica (MURST).
Bibliografia
Alberch, P., 1982. Developmental constraints in evolutionary processes. In: J.T.Bonner ed., Evolution and development, Springer-Verlag, Berlin Heidelberg, pp. 313-332.
Alexander, R.M.C.N., 1985. The ideal and the feasible: physical constraints on evolution. Biol. J. Linn. Soc., 26:: 345-358.
Amundson, R., 1994. Two concepts of constraint. adaptationism and the challenge from developmental biology. Phil Sci., 61: 556-578.
Antonovics, J. e Van Tienderen, P.H., 1991. Ontoecogenophyloconstraints? The chaos of constraint terminology. Tr. Ecol. Evol., 6: 166-167.
Arnold, S.J., 1992. Constraints on phenotypic evolution. Am. Nat., 140 Suppl.: S85-S107.
Avital, E. e Jablonka, E., 1994. Social learning and the evolution of behaviour. Anim. Behav., 48: 1195-1199.
Baldwin, J.M., 1896. A new factor in evolution. Am. Nat., 30: 441-541.
Barbieri, M., 1997. Biological forms as natural conventions. Riv. Biol. /Biol. Forum, 90: 485-488.
Berrill, N.J. e Goodwin, B.C., 1996. The life of form. Emergent patterns of morphological transformation. Riv. Biol. /Biol. Forum, 89: 373-388.
Brink, R.A., 1960. Paramutation and chromosome organization. Q. Rev. Biol., 35: 120-137.
Biürger, R., 1986. Constraints for the evolution of functionally coupled characters: a nonlinear analysis of a phenotypic model. Evolution, 40: 182-193.
Carrier, D.R., 1991. Conflict in the hypaxial musculo-skeletal system: documenting an evolutionary constraint. Am. Zool., 31: 644-654.
Carrière, Y., 1994. Evolution of phenotypic variance: non-Mendelian parental influences on phenotypic and genotypic components of life-history traits in a generalist herbivore. Heredity, 72: 420-430.
Derrickson, E.M. e Ricklefs, R.E., 1988. Taxon-dependent diversification of life-history traits and the perception of phylogenetic constraints. Funct. Ecol., 2: 417-423.
Dover, G., 1982. Molecular drive: a cohesive mode of species evolution. Nature, 299: 111-117.
Dullemeijer, P., 1991. Evolution of biological constructions: concessions, limitations and pathways. In: N. Schmidt-Kittler e V. Vogel eds. Constructional morphology and evolution. Springer-Verlag, Berlin Heidelberg, pp. 313-323.
Dullemeijer, P. e Zweers, G.A., 1997. The variety of explanations of living forms and structures. Eur. J. Morph., 35: 354-364.
Faulkes, C.G., N.C. Bennett, M.W. Bruford, O’Brien, H.P, Aguilar, G.H. e Jarvis, J.U.M., 1997. Ecological constraints drive social evolution in the African mole-rats. Proc. R.Soc London B, 264: 1619-1627.
Gerard, J.F., Laffort, B. e Vancassel, M., 1992. Theoretical possibility of a genetic assimilation of the fleeing direction in Talitrus saltator. Behav. Processes, 27: 195-204.
Gould, S.J., 1980. The evolutionary biology of constraint. Daedalus, 109: 39-52.
Gould, S.J., 1989. A developmental constraint in Cerion, with comments on the definition and interpretation of constraint in evolution. Evolution, 43: 516-539.
Hall, B.K., 1986. The role of movement and tissue interactions in the development and growth of bone and secondary cartilage in the clavicle of the embryonic chick. J. Embryol. Exp. Morphol., 93: 133-152.
Herring, S.W., 1993. Epigenetic and functional influences on skull growth. In: The Skull, vol.1, J.Hanken e B.K. Hall eds., University of Chicago Press, pp. 153-206.
Holliday, R., 1987. The inheritance of epigenetic defects. Science, 238: 163-170.
Ingber, D.E., 1993. Cellular tensegrity, defining new rules of biological design that govern the cytoskeleton. J. Cell Sci., 103.
Ingber, D.E., 1997. Tensegrity: the architectural basis of cellular mechanotransduction. Ann. Rev. Physiol., 59.
Jablonka, E. e Lamb, M.J., 1995. Epigenetic inheritance and evolution: the Lamarckian dimension. Oxford University Press.
Jorgensen, R., 1994. Developmental significance of epigenetic impositions on the plant genome: a paragenetic function for chromosomes. Dev. Genet., 15: 523-532.
Kauffman, S., 1983. Developmental constraints. internal factors in evolution. In B. Goodwin, N. Holder e C.C. Wylie (eds) Development and evolution. Cambridge University Press, Cambridge, pp. 195-225.
Kauffman, S., 1993. The origins of order. Oxford University Press, Oxford.
Kirkpatrick, M e Lande, R., 1989. The evolution of maternal characters. Evolution,43: 485-503.
Lauder, G.V., 1996. The argument from design. I. Adaptation, Ac. Press, pp. 55-91.
Maynard Smith, J., Burian, R., Kauffman, S., Alberch, P., Campbell, J., Goodwin, B., Lande, R., Raup, D. e Wolpert, L., 1985. Developmental constraints and evolution. Q. Rev. Biol., 60: 265-287.
McKitrick, M.C., 1993. Phylogenetic constraint in evolutionary theory: has it any explanatory power? Annu. Rev. Ecol. Syst., 24: 307-330.
Mikula, B. C., 1995. Environmental programming of heritable epigenetic changes in paramutant r-gene expression using temperature and light at a specific stage of early development in maize seedlings. Genetics, 140: 1379-1387.
Monk, M., 1995. Epigenetic programming of differential gene expression in development and evolution. Dev. Genet., 17: 188-197.
Moran, N.A., 1994. Adaptation and constraint in the complex life cycles of animals. Annu. Rev. Ecol. Syst., 25: 573-600.
Moreno, G., 1994. Genetic architecture, genetic behavior, and character evolution. Annu. Rev. Ecol Syst., 25: 31-44.
Müller, G.B. e Wagner, G.P., 1996. Homology, Hox genes, and developmental integration. Am. Zool., 36: 4-13.
Newman, S.A. e Comper, W.D., 1990. "Generic" physical mechanisms of morphogenesis and pattern formation. Development , 110: 1-18.
Pigliucci, M. e Schlichting, C.D., 1997: On the limits of quantitative genetics for the study of phenotypic evolution. Acta Biotheor., 45: 143-160.
Prigogine, I. e Stenger, I., 1997. La fin des certitudes. Edition Odile Jacob, Paris.
Riedl, G., 1978. Order in living organisms. Wiley, New Yprk.
Rossiter, M.C., 1996. Incidence and consequences of inherited environmental effects. Annu. Rev. Ecol. Syst., 27: 451-476.
Sarà, M., 1993. Biological evolution: an holistic organism-centered approach. Riv. Biol./Biol. Forum, 86: 347-359.
Smith-Gill, S.J., 1983. Developmental plasticity: developmental conversion versus phenotypic modulation. Amer. Zool., 23: 47-55.
Stearns, S.C., 1986. Natural selection and fitness, adaptation and constraint.: In: D.M.Raup e D.Jablonski (eds.) Patterns and processes in the history of life. Springer-Verlag, New York.
Stephens, D.W. e Krebs, J.R., 1986: Foraging theory. Princeton University Press, Princeton.
Sternberg, R. von, 1996. Genome self-modification and cellular control of genome reorganization. Riv. Biol./ Biol. Forum, 89: 423-462.
Sternberg, R.M. von, Novick, G.M., Gao, G.P. e Herrera, R.J., 1992. Genome canalization: the coevolution of transposable and interspersed repetitive elements with single copy DNA. Genetica, 86: 215-246.
Tierney, A.J., 1986. The evolution of learned and innate behavior: contributions from genetics and neurobiology to a theory of behavioral evolution. Anim. Learn. Behav., 14: 339-348.
Vancassel, M., 1990. Behavioural development and adaptation: an assimilation of some of Waddington’s ideas. Behav. Processes, 22: 23-31.
Van Essen, D. C., 1997. A tension-based theory of morphogenesis and compact wiring in the central nervous system. Nature, 385: 313-318.
Waddington, C.H., 1957. The strategy of the genes. Allen and Unwin, London.
Waddington, C.H., 1975. The evolution of an evolutionist. Edinburgh Univ. Press, Edinburgh.
Wagner, G.P., 1988. The significance of developmental constraints for phenotypic evolution by natural selection. In: G. de Jong (ed.) Population genetics and evolution, Springer-Verlag, Berlin Heidelberg, pp. 222-229.
West-Eberhard, M.J., 1989. Phenotypic plasticity and the origins of diversity. Annu. Rev. Ecol. Syst., 20: 249-278.
Wolf, J.B., Brodie III, E.D., Cheverud, J.M., Moore, A.J. e Wade, M.J., 1998: Evolutionary consequences of indirect genetic effects. Tr. Ecol. Evol., 13: 64-69.